- Tällä geenillä on muita nimiä SPRING ja RNF91.
- Sen rakenne on RBCC (RING, Bbox1, Bbox2, Coiled-coil) ja
C-terminaalin osalta alaluokkaa C_I (FN3, PRY-SPRY).
- Geeni koodaa kahta isoformia. Toisessa, pidemmässä, on 710
aminohappoa (isoformi1). Domeenien sijainnit ovat :
RING( 9..38),
Bbox (224..266), Bbox ( 273..398). FN3 domeenit sijoittuvat jaksoon
453..532. SPRY..PRY jakso on 529..700 ja se on samankaltainen myös
TRIM67:ssä, joka myös kuuluu tähän alaluokaan C-I .
FN3- domeeni
tarkoittaa fibronektiini tyyppi3:n kaltaista domeenia.
- TRIM9 proteiini lokalisoituu sytoplasmisiin kappaleisiin. Sen
funktiota ei ollut tunnistettu vielä 2008 mennessä, mutta paljon
artikkelia on kertynyt sen jälkeen. Geeniä ilmenee eniten
aivoissa, sappirakossa ja vielä hieman yhdessä kudoksessa.
Alla on PubMed Gene lähteestä tiedot 1-isoformista, joka on pitempi muoto. Lyhemmästä mudosota on artikkeli, jok selvittää TRIM).n duaalivaikutuksista osuutta antiviraalissa immuunivasteessa. Aivoissa netriini- 1 vaikuttaa olevan tärkeä aksonin kunnolle ja sen signaalijärjestelmään TRIM9 vuorovaikutaa, muta mekanismi ei ole aivan yksinkertaisen selkeä, eikä näihin selityksiin liitetä trimperheen keskeisiä vuorovaikutuksia.
- https://www.ncbi.nlm.nih.gov/gene/114088
-
- RNF91; SPRING Summary.The protein encoded by this gene is a member
of the tripartite motif (TRIM) family. The TRIM motif includes three
zinc-binding domains, a RING, a B-box type 1 and a B-box type 2, and
a coiled-coil region. The protein localizes to cytoplasmic bodies.
Its function has not been identified. Alternate splicing of this
gene generates two transcript variants encoding different isoforms.
[provided by RefSeq, Jul 2008] Expression Biased expression in
brain (RPKM 15.3), gall bladder (RPKM 1.4) and 1 other tissue See
more
Mielenkiintoisia artikkeleita TRIM9 funktioista netriini-1
säätelyjärjestelmässä
Avainsanoja:
DCC tässä
yhteydessä tarkoittaa ”deleted in colorectal cancer”, ”puuttuu
kolorektaalisyövässä”. DCC on netriini-1 reseptori.
KUVA:
(Suomennosta):
TRIM9:stä riippuva
netriini1-reseptorin (DDC) ubikitinaatio pitää aisoissa
kinaasisignalointia, exosytoosia ja aksonien haaroittumista.
Extrasellulaarinen netriini-1 ja sen reseptori DCC edistävät
aivokuoren kehittyvässä neuronissa neuroninjatkeen (aksonin)
haaroittumista. Netriinistä riippuvaa morfogeneesiä edeltää
netriinireseptorin (DCC) multimerisoituminen, Kinaasin
FAK-aktivaatio js Src-perheen kinaasien aktivoituminen – ja se
lisää exosyyttisten rakkuloiden fuusioitumista- mutta näiden
tapahtumien keskinäinen linkkiytyminen on ollut tuntematon seikka.
Tässä työssä
tutkijat osoittavat, että TRIM9:n tekemä netriinireseptorin (DCC)
ubikitinaatio estää FAK- interaktion ja fosforyloitumisen. Mutta
kun netriini-1 stimuloi tätä reseptoriaan DCC, niin TRIM9 edistää
reseptorin multimerisaatiota, mutta samalla vähenee DCC:n
ubikitinoitumisen aste, mistä taas fokaaliadheesiokinaasin (FAK)
interaktio pääsee edistymään ja samoin sen aktivaatio.
Tutkijat
havaitsivat, että FAK-aktiivisuuden inhibitio ( estäminen)
blokeerasi exosytoosin kohonneet frekvenssit koeputkessa ( in
vitro) ja aksonihaaroittumisen lisääntymisen in vitro ja in
vivo.
Vaikka
fokaaliadheesiokinaasin vähensi SNARE- välitteistä exosytoosia,
SNARE-kompleksien ja rakkuloiden kokoontuminen plasmakalvon
läheisyyteen lisääntyi , mikä viittaisi siihen, että FAK omaa
jonkin uuden roolin SNARE-kompleksien kokoontumisen progressiossa
ja rakkuloiden fuusioitumisessa koe-eläimen kehittyvässä
neuronissa.
-
TRIM9-dependent
ubiquitination of DCC constrains kinase signaling, exocytosis, and
axon branching. Extracellular netrin-1 and its receptor deleted
in colorectal cancer (DCC) promote axon branching in developing
cortical neurons. Netrin-dependent morphogenesis is preceded by
multimerization of DCC, activation of FAK and Src family kinases,
and increases in exocytic vesicle fusion, yet how these occurrences
are linked is unknown.
-
Here we
demonstrate that tripartite motif protein 9
(TRIM9)-dependent ubiquitination of DCC blocks the interaction with
and phosphorylation of FAK. Upon netrin-1 stimulation TRIM9 promotes
DCC multimerization, but TRIM9-dependent ubiquitination of DCC is
reduced, which promotes an interaction with FAK and subsequent FAK
activation.
-
We found
that inhibition of FAK activity blocks elevated frequencies of
exocytosis in vitro and elevated axon branching in vitro and in
vivo. Although FAK inhibition decreased soluble
N-ethylmaleimide attachment protein receptor
(SNARE)-mediated exocytosis, assembled SNARE complexes and vesicles
adjacent to the plasma membrane increased, suggesting a novel role
for FAK in the progression from assembled SNARE complexes to vesicle
fusion in developing murine neurons.
Toinen artikkeli ja kuva netriini-1
signaloinnista ja TRIM9 osuudesta
Tunnettuja TRIM9-
interaktioita.
Sanastoa: F-actin,
on filamenttimuotoinen aktiini.
Kuva A.
Netriini-1:n poissaollessa TRIM9 inhiboi SNARE kompleksin jäseniä
(kuten SNAP25) ligaasifunktiostaan riippumattomalla tavalla estäen
rakkuloiden exosytoosia. TRIM9 toimii filopodia rakenteille ”OFF”
vaihteen tavoin ubikitinoimalla VASP-polymeraasin. Ubikitinoitu
prosessiivinen aktiinipolymeraasi VASP irtoaa aktiinifilamenttien
päistä jolloin aktiinipolymerisaatio vähenee.
Kuva B. Kun
netriini-1 on läsnä, TRIM9 tekee interaktion netriini-1-reseptorin
(DCC) kanssa ligaasista riippuvalla tavalla. TRIM9:n interaktio
SNAP25:n kanssa estyy, kun netriini-1 tulee reseptoriinsa. Ja nyt
pääsee tapahtumaan exosytoosivälitteistä kalvon laajenemista ja
filopodia ”ON” vaihde, sillä TRIM9:n tekemä VASP-ubikitinaatio
estyy ja VASP ( aktiinipolymeraasi ) pääsee assosioitumaan
aktiinifilamentteihin ja aktiinifilamentit alkavat polymerisoitua ja
filopodiamuodostus edistyy.
-
Figure 1.
Known TRIM9 interactions. A, In the absence of Netrin-1, TRIM9
inhibits SNAP25 in a ligase-independent manner to inhibit
exocytosis. TRIM9 also acts as a filopodia " off " switch
by ubiquitinating actin polymerase VASP. Ubiquitinated VASP is
dissociated from the ends of actin filaments, leading to decreased
actin polymerization. B, When Netrin-1 is present, TRIM9 interacts
with Netrin-1 receptor DCC in a ligase-dependent manner. TRIM9's
interaction with SNAP25 is inhibited upon Netrin-1 introduction,
leading to exocytosis-mediated membrane expansion. TRIM9
ubiquitination of VASP is also inhibited, leading to VASP
association with actin filaments, increased actin filament
polymerization, and promotion of filopodia formation. F-actin,
filamentous actin; Ub, ubiquitination.
Cell
Res. 2016 May;26(5):613-28. doi: 10.1038/cr.2016.27. Epub 2016
Feb 26.
Lyhempi TRIM9 isoformi omaa kaksoisvaikutuksia
antivirusimmuunivasteessa.
-
TRIM9 short
isoform preferentially promotes DNA and RNA virus-induced production
of type I interferon by recruiting GSK3β to TBK1.
Qin
Y1,
Liu
Q1,
Tian
S1,
Xie
W2,
Cui
J1,3,
Wang
RF4,5,6.
(Suomennosta) .
Avainsanoja:
MAVS =Mitochondrial
Anti-Viral Signaling-protein
STING=double-strand
(ds) DNA receptors activate the adaptor molecule STimulator of
IFN Genes (STING) at the endoplasmic reticulum (ER)
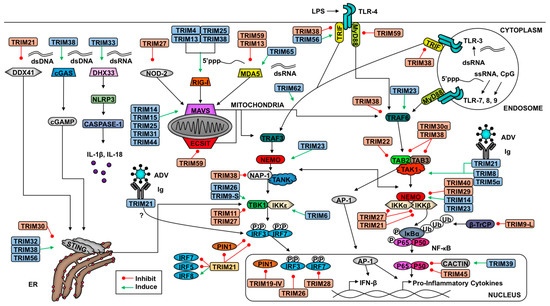
Kuva tässä
linkissä osoittaa MAVS ja STING proteiinien sijainnin. MAVS on
mitokondriaan assosioitunut ja STING endoplasmiseen retikulumiin.
Niitten avulla solu lajittelee virusvihollisiaan.
”
Antivirusimmuunivasteessa on tyypin 1 interferoni(IFN)
signaalinvälittäjäna ja sen sanomana on, että solun
virussensorijärjestelmä (DNA-sensori cGAS ja RNA-sensorit RIG-1
ja MDA5) ovat tunnistaneet DNA ja RNA virusta. Näiden DNA ja
RNA-sensoreiden aktivoituminen johtaa STING- ja vastaavasti MAVS-
proteiinien rekrytoitumiseen- ja nämä herätteet konvergoituvat
TBK1- signalointitiehen ja siitä seuraa IRF3- fosforyloituminen (
ja aktivoituminen) ja 1 tyypin IFN:n tuotto ( katso kuva)
-
Type I
interferon (IFN) is an important component of antiviral innate
immune signaling mediated by viral DNA and RNA recognition by the
DNA sensor cGAS and RNA sensors RIG-I and MDA5. Activation of these
DNA and RNA sensors leads to the recruitment of STING and MAVS,
respectively, and converges on TANK-binding kinase 1 (TBK1)
signaling for subsequent phosphorylation of IFN regulatory factor 3
(IRF3).
(TANK= TRAF family
member associated NfkB activator)
(TRAF = TNR receptor
associated factor)
(TNR = tumor
necrosis factor, proinflamamtorinen sytokiini)
(
GSK3beta
=glykogeenisyntaasikinaasi , 3 beta
https://en.wikipedia.org/wiki/GSK-3,
kiinnostava entsyymi ja ilmenee monessa patologiassa. Pitää katsoa aivan erikseen sen osuus taudeissa , se pulpahtaa esille mitä erilaisimmissa yhteyksissä).
”TBK1 kinaasin
aktivaatio ei ole täysin yksityiskohtaisesti selvitetty. Tässä
työssä on tunnistettu TRIM9 geenin koodaama lyhempi isoformi
1-tyypin interferonisignaloinnin positiiviseksi säätelijäksi.
Virusinfektion aikana TRIM9 autopolyubikitinoituu K63 asemaan ja
toimii alustana, jolla GSK3beta liittyy TBK1 entsyymiin ja tätä
tietä pääsee IRF3-signalointi aktivoitumaan. Tutkijat
havaitsivat, että TRIM9 pystyy selektiivisesti inhiboimaan
proinflammatoristen sytokiinien tuotantoa, mutta lisää 1- tyypin
interferonin ja interferonilla stimuloituvien geenien ilmenemistä.
Tutkijoitten löytönä on TRIM9:n kaksoisvaikutukset
antivirusimmuniteetissä ja ne tasapainottavat proinflammatorisia
vasteita ja 1- tyypin interferonien tuotantoa”.
-
However, the
mechanisms that control TBK1 activation are still poorly defined.
Here, we identify tripartite motif 9 short isoform (TRIM9s) as a
positive regulator in type I IFN signaling. Upon viral infection,
TRIM9s undergoes Lys-63-linked auto-polyubiquitination and serves as
a platform to bridge GSK3β to TBK1, leading to the activation of
IRF3 signaling. Interestingly, we found that TRIM9s selectively
inhibits the production of pro-inflammatory cytokines, but enhances
the expression of type I IFNs as well as IFN-stimulated genes, in
response to viral infection. Our findings reveal novel dual
functions of TRIM9s in antiviral immunity, which serve to balance
pro-inflammatory response and production of type I IFNs.
-
TRIM9 on aivospesifinen NF-kB:n negatiivinen
säätelijä.
- TRIM9 as a brain-specific negative
regulator of the NF-κB pro-inflammatory signalling pathway.
TRIM9 pitempi isoformi 1
E3 ubiquitin-protein
ligase TRIM9 isoform 1 [Homo sapiens]
NCBI Reference Sequence: NP_055978.4
Identical
Proteins FASTA
Graphics
LOCUS NP_055978 710 aa linear PRI 08-APR-2018
DEFINITION E3 ubiquitin-protein ligase TRIM9 isoform 1 [Homo sapiens].
ACCESSION NP_055978
VERSION NP_055978.4
DBSOURCE REFSEQ: accession NM_015163.5
KEYWORDS RefSeq.
SOURCE Homo sapiens (human)
ORGANISM Homo sapiens
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi;
Mammalia; Eutheria; Euarchontoglires; Primates; Haplorrhini;
Catarrhini; Hominidae; Homo.
REFERENCE 1 (residues 1 to 710)
AUTHORS Plooster M, Menon S, Winkle CC, Urbina FL, Monkiewicz C, Phend KD,
Weinberg RJ and Gupton SL.
TITLE TRIM9-dependent ubiquitination of DCC constrains kinase signaling,
exocytosis, and axon branching Extracellular netrin-1 and its receptor deleted in colorectal cancer (DCC) promote axon branching in developing cortical neurons.
JOURNAL Mol. Biol. Cell 28 (18), 2374-2385 (2017)
PUBMED 28701345
REMARK GeneRIF: Authors demonstrate that tripartite motif protein 9
(TRIM9)-dependent ubiquitination of DCC blocks the interaction with
and phosphorylation of FAK.
REFERENCE 2 (residues 1 to 710)
AUTHORS Qin Y, Liu Q, Tian S, Xie W, Cui J and Wang RF.
TITLE TRIM9 short isoform preferentially promotes DNA and RNA
virus-induced production of type I interferon by recruiting
GSK3beta to TBK1
JOURNAL Cell Res. 26 (5), 613-628 (2016)
PUBMED 26915459
REMARK GeneRIF: TRIM9s undergoes Lys-63-linked auto-polyubiquitination and
serves as a platform to bridge GSK3beta to TBK1, leading to the
activation of IRF3 signaling.
REFERENCE 3 (residues 1 to 710)
AUTHORS Yang X, Coulombe-Huntington J, Kang S, Sheynkman GM, Hao T,
Richardson A, Sun S, Yang F, Shen YA, Murray RR, Spirohn K, Begg
BE, Duran-Frigola M, MacWilliams A, Pevzner SJ, Zhong Q, Trigg SA,
Tam S, Ghamsari L, Sahni N, Yi S, Rodriguez MD, Balcha D, Tan G,
Costanzo M, Andrews B, Boone C, Zhou XJ, Salehi-Ashtiani K,
Charloteaux B, Chen AA, Calderwood MA, Aloy P, Roth FP, Hill DE,
Iakoucheva LM, Xia Y and Vidal M.
TITLE Widespread Expansion of Protein Interaction Capabilities by
Alternative Splicing
JOURNAL Cell 164 (4), 805-817 (2016)
PUBMED 26871637
REFERENCE 4 (residues 1 to 710)
AUTHORS Shi M, Cho H, Inn KS, Yang A, Zhao Z, Liang Q, Versteeg GA,
Amini-Bavil-Olyaee S, Wong LY, Zlokovic BV, Park HS, Garcia-Sastre
A and Jung JU.
TITLE Negative regulation of NF-kappaB activity by brain-specific
TRIpartite Motif protein 9
JOURNAL Nat Commun 5, 4820 (2014)
PUBMED 25190485
REMARK GeneRIF: TRIM9 is a brain-specific negative regulator of the
NF-kappaB pro-inflammatory signalling pathway.
Publication Status: Online-Only
REFERENCE 5 (residues 1 to 710)
AUTHORS Kanazawa T, Ikeda M, Glatt SJ, Tsutsumi A, Kikuyama H, Kawamura Y,
Nishida N, Miyagawa T, Hashimoto R, Takeda M, Sasaki T, Tokunaga K,
Koh J, Iwata N and Yoneda H.
TITLE Genome-wide association study of atypical psychosis
JOURNAL Am. J. Med. Genet. B Neuropsychiatr. Genet. 162B (7), 679-686
(2013)
PUBMED 24132900
REFERENCE 6 (residues 1 to 710)
AUTHORS Tanji K, Kamitani T, Mori F, Kakita A, Takahashi H and Wakabayashi
K.
TITLE TRIM9, a novel brain-specific E3 ubiquitin ligase, is repressed in
the brain of Parkinson's disease and dementia with Lewy bodies
JOURNAL Neurobiol. Dis. 38 (2), 210-218 (2010)
PUBMED 20085810
REMARK GeneRIF: These results suggest that TRIM9 plays an important role
in the regulation of neuronal functions and participates in
pathological process of Lewy body disease through its ligase
activity.
REFERENCE 7 (residues 1 to 710)
AUTHORS Li Y, Chin LS, Weigel C and Li L.
TITLE Spring, a novel RING finger protein that regulates synaptic vesicle
exocytosis
JOURNAL J. Biol. Chem. 276 (44), 40824-40833 (2001)
PUBMED 11524423
REFERENCE 8 (residues 1 to 710)
AUTHORS Reymond A, Meroni G, Fantozzi A, Merla G, Cairo S, Luzi L,
Riganelli D, Zanaria E, Messali S, Cainarca S, Guffanti A, Minucci
S, Pelicci PG and Ballabio A.
TITLE The tripartite motif family identifies cell compartments
JOURNAL EMBO J. 20 (9), 2140-2151 (2001)
PUBMED 11331580
REFERENCE 9 (residues 1 to 710)
AUTHORS Ohara O, Nagase T, Ishikawa K, Nakajima D, Ohira M, Seki N and
Nomura N.
TITLE Construction and characterization of human brain cDNA libraries
suitable for analysis of cDNA clones encoding relatively large
proteins
JOURNAL DNA Res. 4 (1), 53-59 (1997)
PUBMED 9179496
REFERENCE 10 (residues 1 to 710)
AUTHORS Dawson SJ and White LA.
TITLE Treatment of Haemophilus aphrophilus endocarditis with
ciprofloxacin
JOURNAL J. Infect. 24 (3), 317-320 (1992)
PUBMED 1602151
COMMENT REVIEWED REFSEQ: This record has been curated by NCBI staff. The
reference sequence was derived from BC063872.1, DA101991.1,
DA173440.1 and D87458.2.
On Jun 13, 2008 this sequence version replaced NP_055978.3.
Summary: The protein encoded by this gene is a member of the
tripartite motif (TRIM) family. The TRIM motif includes three
zinc-binding domains, a RING, a B-box type 1 and a B-box type 2,
and a coiled-coil region. The protein localizes to cytoplasmic
bodies. Its function has not been identified. Alternate splicing of
this gene generates two transcript variants encoding different
isoforms. [provided by RefSeq, Jul 2008].
Transcript Variant: This variant (1) represents the longest
transcript and encodes the longer isoform (1).
Publication Note: This RefSeq record includes a subset of the
publications that are available for this gene. Please see the Gene
record to access additional publications.
##Evidence-Data-START##
Transcript exon combination :: BC063872.1, SRR1660807.103429.1
[ECO:0000332]
RNAseq introns :: mixed/partial sample support
SAMEA1965299, SAMEA1966682
[ECO:0000350]
##Evidence-Data-END##
FEATURES Location/Qualifiers
source 1..710
/organism="Homo sapiens"
/db_xref="taxon:9606"
/chromosome="14"
/map="14q22.1"
Protein 1..710
/product="E3 ubiquitin-protein ligase TRIM9 isoform 1"
/EC_number="2.3.2.27"
/note="homolog of rat RING finger Spring; tripartite
motif-containing protein 9; E3 ubiquitin-protein ligase
TRIM9; RING finger protein 91; RING-type E3 ubiquitin
transferase TRIM9"
/calculated_mol_wt=79046
Region 9..>38
/region_name="RING"
/note="RING-finger (Really Interesting New Gene) domain, a
specialized type of Zn-finger of 40 to 60 residues that
binds two atoms of zinc; defined by the 'cross-brace'
motif C-X2-C-X(9-39)-C-X(1-3)-
H-X(2-3)-(N/C/H)-X2-C-X(4-48)C-X2-C; probably involved
in...; cd00162"
/db_xref="CDD:238093"
Site 41
/site_type="phosphorylation"
/experiment="experimental evidence, no additional details
recorded"
/note="Phosphothreonine. {ECO:0000250|UniProtKB:Q8C7M3};
propagated from UniProtKB/Swiss-Prot (Q9C026.1)"
Site 44
/site_type="phosphorylation"
/experiment="experimental evidence, no additional details
recorded"
/note="Phosphoserine. {ECO:0000250|UniProtKB:Q8C7M3};
propagated from UniProtKB/Swiss-Prot (Q9C026.1)"
Site 46
/site_type="phosphorylation"
/experiment="experimental evidence, no additional details
recorded"
/note="Phosphoserine. {ECO:0000250|UniProtKB:Q8C7M3};
propagated from UniProtKB/Swiss-Prot (Q9C026.1)"
Site 49
/site_type="phosphorylation"
/experiment="experimental evidence, no additional details
recorded"
/note="Phosphoserine. {ECO:0000250|UniProtKB:Q8C7M3};
propagated from UniProtKB/Swiss-Prot (Q9C026.1)"
Region 163..211
/region_name="BBOX"
/note="B-Box-type zinc finger; smart00336"
/db_xref="CDD:197662"
Region 224..266
/region_name="BBOX"
/note="B-Box-type zinc finger; smart00336"
/db_xref="CDD:197662"
Region 273..399
/region_name="BBC"
/note="B-Box C-terminal domain; smart00502"
/db_xref="CDD:128778"
Region 453..532
/region_name="FN3"
/note="Fibronectin type 3 domain; One of three types of
internal repeats found in the plasma protein fibronectin.
Its tenth fibronectin type III repeat contains an RGD cell
recognition sequence in a flexible loop between 2 strands.
Approximately 2% of all...; cd00063"
/db_xref="CDD:238020"
Site order(521..522,524..525)
/site_type="other"
/note="Cytokine receptor motif"
/db_xref="CDD:238020"
Region 529..700
/region_name="SPRY_PRY_TRIM67_9"
/note="PRY/SPRY domain in tripartite motif-containing
proteins, TRIM9 and TRIM67; cd12889"
/db_xref="CDD:293947"
CDS 1..710
/gene="TRIM9"
/gene_synonym="RNF91; SPRING"
/coded_by="NM_015163.5:766..2898"
/note="isoform 1 is encoded by transcript variant 1"
/db_xref="CCDS:CCDS9703.1"
/db_xref="GeneID:114088"
/db_xref="HGNC:HGNC:16288"
/db_xref="MIM:606555"
ORIGIN
1 meemeeelkc pvcgsfyrep iilpcshnlc qacarnilvq tpesespqsh raagsgvsdy
61 dyldldkmsl yseadsgygs yggfasaptt pcqkspngvr vfppampppa thlspalapv
121 prnscitcpq chrslilddr glrgfpknrv legvidryqq skaaalkcql cekapkeatv
181 mceqcdvfyc dpcrlrchpp rgplakhrlv ppaqgrvsrr lsprkvstct dhelenhsmy
241 cvqckmpvcy qcleegkhss hevkalgamw klhksqlsqa lnglsdrake akeflvqlrn
301 mvqqiqensv efeaclvaqc dalidalnrr kaqllarvnk ehehklkvvr dqishctvkl
361 rqttglmeyc levikendps gflqisdali rrvhltedqw gkgtltprmt tdfdlsldns
421 pllqsihqld fvqvkasspv patpilqlee ccthnnsatl swkqpplstv padgyileld
481 dgnggqfrev yvgketmctv dglhfnstyn arvkafnktg vspysktlvl qtsevawfaf
541 dpgsahsdii lsndnltvtc ssyddrvvlg ktgfskgihy weltvdrydn hpdpafgvar
601 mdvmkdvmlg kddkawamyv dnnrswfmhn nshtnrtegg itkgatigvl ldlnrknltf
661 findeqqgpi afdnveglff pavslnrnvq vtlhtglpvp dfyssrasia
Search results
Items: 1 to 20 of 25
<< First< Prev
Page of 2
Select item 290213671.
Tokarz DA, Heffelfinger AK, Jima DD, Gerlach J, Shah RN,
Rodriguez-Nunez I, Kortum AN, Fletcher AA, Nordone SK, Law JM, Heber
S, Yoder JA.
J Leukoc Biol. 2017 Dec;102(6):1371-1380. doi:
10.1189/jlb.1A0816-371R. Epub 2017 Oct 11.
- The vertebrate immune response comprises multiple molecular and
cellular components that interface to provide defense against
pathogens. Because of the dynamic complexity of the immune system
and its interdependent innate and adaptive functionality, an
understanding of the whole-organism response to pathogen exposure
remains unresolved. Zebrafish larvae provide a unique model for
overcoming this obstacle, because larvae are protected against
pathogens while lacking a functional adaptive immune system during
the first few weeks of life. Zebrafish larvae were exposed to immune
agonists for various lengths of time, and a microarray transcriptome
analysis was executed. This strategy identified known immune
response genes, as well as genes with unknown immune function,
including the E3 ubiquitin ligase tripartite motif-9 (Trim9).
Although trim9 expression was originally described as "brain
specific," its expression has been reported in stimulated human
Mϕs. In this study, we found elevated levels of trim9
transcripts in vivo in zebrafish Mϕs after immune stimulation.
Trim9 has been implicated in axonal migration, and we therefore
investigated the impact of Trim9 disruption on Mϕ motility and
found that Mϕ chemotaxis and cellular architecture are subsequently
impaired in vivo. These results demonstrate that Trim9 mediates
cellular movement and migration in Mϕs as well as neurons.
Similar
articles
Select item 287013452.
Plooster M, Menon S, Winkle CC, Urbina FL, Monkiewicz C, Phend KD,
Weinberg RJ, Gupton SL.
Mol Biol Cell. 2017 Sep 1;28(18):2374-2385. doi:
10.1091/mbc.E16-08-0594. Epub 2017 Jul 12.
Abstract
Extracellular netrin-1 and its receptor deleted in colorectal
cancer (DCC) promote axon branching in developing cortical neurons.
Netrin-dependent morphogenesis is preceded by multimerization of DCC,
activation of FAK and Src family kinases, and increases in exocytic
vesicle fusion, yet how these occurrences are linked is unknown. Here
we demonstrate that
tripartite
motif protein 9
(TRIM9)-dependent ubiquitination of DCC blocks the interaction with
and phosphorylation of FAK. Upon netrin-1 stimulation TRIM9 promotes
DCC multimerization, but TRIM9-dependent ubiquitination of DCC is
reduced, which promotes an interaction with FAK and subsequent FAK
activation. We found that inhibition of FAK activity blocks elevated
frequencies of exocytosis in vitro and elevated axon branching in
vitro and in vivo. Although FAK inhibition decreased
soluble
N-ethylmaleimide
attachment protein
receptor
(SNARE)-mediated exocytosis, assembled SNARE complexes and vesicles
adjacent to the plasma membrane increased, suggesting a novel role
for FAK in the progression from assembled SNARE complexes to vesicle
fusion in developing murine neurons.
Free PMC
Article
Similar
articles
Select item 285617363.
Nagarkar-Jaiswal S, Manivannan SN, Zuo Z, Bellen HJ.
Elife. 2017 May 31;6. pii: e26420. doi: 10.7554/eLife.26420.
- PMID:
- 28561736
Free PMC
Article
Similar
articles
Select item 277434774.
Akin O, Zipursky SL.
Elife. 2016 Oct 15;5. pii: e20762. doi: 10.7554/eLife.20762.
- PMID:
- 27743477
Free PMC
Article
Similar
articles
Select item 276297035.
Aiken J, Buscaglia G.
J Neurosci. 2016 Sep 14;36(37):9513-5. doi:
10.1523/JNEUROSCI.1917-16.2016. No abstract available.
- PMID:
- 27629703
Free Article
Similar
articles
Select item 273932366.
Liu Y, Li J, Wang F, Mao F, Zhang Y, Zhang Y, Yu Z.
Fish Shellfish Immunol. 2016 Sep;56:106-110. doi:
10.1016/j.fsi.2016.06.057. Epub 2016 Jul 5.
- PMID:
- 27393236
Similar
articles
Select item 271476497.
Winkle CC, Olsen RH, Kim H, Moy SS, Song J, Gupton SL.
J Neurosci. 2016 May 4;36(18):4940-58. doi:
10.1523/JNEUROSCI.3876-15.2016.
- PMID:
- 27147649
Free PMC
Article
Similar
articles
Select item 269154598.
Qin Y, Liu Q, Tian S, Xie W, Cui J, Wang RF.
Cell Res. 2016 May;26(5):613-28. doi: 10.1038/cr.2016.27. Epub
2016 Feb 26.
- PMID:
- 26915459
Free PMC
Article
Similar
articles
Select item 267028299.
Menon S, Boyer NP, Winkle CC, McClain LM, Hanlin CC, Pandey D,
Rothenfußer S, Taylor AM, Gupton SL.
Dev Cell. 2015 Dec 21;35(6):698-712. doi:
10.1016/j.devcel.2015.11.022.
- PMID:
- 26702829
Free PMC
Article
Similar
articles
Select item 2659908210.
Zhan W, Han T, Zhang C, Xie C, Gan M, Deng K, Fu M, Wang JB.
PLoS One. 2015 Nov 24;10(11):e0142596. doi:
10.1371/journal.pone.0142596. eCollection 2015.
- PMID:
- 26599082
Free PMC
Article
Similar
articles
Select item 2654376911.
Mishima C, Kagara N, Matsui S, Tanei T, Naoi Y, Shimoda M,
Shimomura A, Shimazu K, Kim SJ, Noguchi S.
Springerplus. 2015 Oct 22;4:635. doi: 10.1186/s40064-015-1423-7.
eCollection 2015.
- PMID:
- 26543769
Free PMC
Article
Similar
articles
Select item 2519048512.
Shi M, Cho H, Inn KS, Yang A, Zhao Z, Liang Q, Versteeg GA,
Amini-Bavil-Olyaee S, Wong LY, Zlokovic BV, Park HS, García-Sastre
A, Jung JU.
Nat Commun. 2014 Sep 5;5:4820. doi: 10.1038/ncomms5820.
- PMID:
- 25190485
Free PMC
Article
Similar
articles
Select item 2477831213.
Winkle CC, McClain LM, Valtschanoff JG, Park CS, Maglione C,
Gupton SL.
J Cell Biol. 2014 Apr 28;205(2):217-32. doi:
10.1083/jcb.201311003.
- PMID:
- 24778312
Free PMC
Article
Similar
articles
Select item 2474679314.
Yang L, Li R, Kaneko T, Takle K, Morikawa RK, Essex L, Wang X,
Zhou J, Emoto K, Xiang Y, Ye B.
Curr Biol. 2014 May 5;24(9):1024-30. doi:
10.1016/j.cub.2014.03.041. Epub 2014 Apr 17.
- PMID:
- 24746793
Free PMC
Article
Similar
articles
Select item 2413290015.
Kanazawa T, Ikeda M, Glatt SJ, Tsutsumi A, Kikuyama H, Kawamura Y,
Nishida N, Miyagawa T, Hashimoto R, Takeda M, Sasaki T, Tokunaga K,
Koh J, Iwata N, Yoneda H.
Am J Med Genet B Neuropsychiatr Genet. 2013 Oct;162B(7):679-86.
doi: 10.1002/ajmg.b.32164.
- PMID:
- 24132900
Similar
articles
Select item 2348276316.
Okaichi K, Izumi N, Takamura Y, Fukui S, Kudo T.
Anticancer Res. 2013 Mar;33(3):923-8.
- PMID:
- 23482763
Similar
articles
Select item 2114318817.
Napolitano LM, Jaffray EG, Hay RT, Meroni G.
Biochem J. 2011 Mar 1;434(2):309-19. doi: 10.1042/BJ20101487.
- PMID:
- 21143188
Similar
articles
Select item 2062707718.
Hao JC, Adler CE, Mebane L, Gertler FB, Bargmann CI,
Tessier-Lavigne M.
Dev Cell. 2010 Jun 15;18(6):950-60. doi:
10.1016/j.devcel.2010.02.019.
- PMID:
- 20627077
Free PMC
Article
Similar
articles
Select item 2008581019.
Tanji K, Kamitani T, Mori F, Kakita A, Takahashi H, Wakabayashi K.
Neurobiol Dis. 2010 May;38(2):210-8. doi:
10.1016/j.nbd.2010.01.007. Epub 2010 Jan 18.
- PMID:
- 20085810
Free PMC
Article
Similar
articles
Select item 1929005320.
Carthagena L, Bergamaschi A, Luna JM, David A, Uchil PD,
Margottin-Goguet F, Mothes W, Hazan U, Transy C, Pancino G, Nisole S.
PLoS One. 2009;4(3):e4894. doi: 10.1371/journal.pone.0004894. Epub
2009 Mar 17.
- PMID:
- 19290053
Free PMC
Article
Similar
articles
Spring, a novel RING finger protein that
regulates synaptic vesicle exocytosis.
Li
Y1,
Chin
LS,
Weigel
C,
Li
L.
Abstract
The synaptosome-associated protein of 25 kDa (SNAP-25) interacts
with syntaxin 1 and vesicle-associated membrane protein 2 (VAMP2) to
form a ternary soluble N-ethylmaleimide-sensitive fusion protein
attachment protein receptor (SNARE) complex that is essential for
synaptic vesicle exocytosis. We report a novel RING finger protein,
Spring, that specifically interacts with SNAP-25. Spring is
exclusively expressed in brain and is concentrated at synapses. The
association of Spring with SNAP-25 abolishes the ability of SNAP-25
to interact with syntaxin 1 and VAMP2 and prevents the assembly of
the SNARE complex. Overexpression of Spring or its
SNAP-25-interacting domain reduces Ca(2+)-dependent exocytosis from
PC12 cells. These results indicate that Spring may act as a regulator
of synaptic vesicle exocytosis by controlling the availability of
SNAP-25 for the SNARE complex formation.
- PMID:
- 11524423
-
DOI:
- 10.1074/jbc.M106141200
- [Indexed for MEDLINE]
Free full text

{kind=link}

{kind=link}
{kind=link}